
Inside every onion cell, thousands of microscopic tunnels are making decisions that will determine how fast the plant grows, how it handles stress, and what kind of organism it becomes. The story begins with a single choice made before the plant even exists: seed or bulb?
A Vegetable With Two Personalities
Picture two onion plants growing side by side in a field. They are the same species, planted in the same soil, watered the same way, exposed to the same sun and rain. By the time harvest comes, they may look nearly identical.
But spend time with them at the molecular level – inside their cells, along their membranes, deep in the activity of their genes – and you find two fundamentally different organisms. One is a sprinter. The other is a strategist. One throws its resources into speed and growth. The other holds back, stays alert, and prioritizes survival.
What created this difference? Not genetics, not environment. Just one thing: where they came from.
One of these plants grew from a seed. The other grew from a small pre-grown bulb called an “onion set.” That single fact – seed or set – triggered a cascade of molecular decisions that rippled through every cell in the plant, reshaping its internal machinery, reconfiguring its plumbing, and setting it on an entirely different biological path.
Recent research has decoded exactly how this happens, and the answer involves a remarkable family of proteins that most people have never heard of. They’re called aquaporins. And once you understand them, you’ll never look at an onion the same way again.
The Plant’s Secret Plumbing
Every cell in every living thing is surrounded by a membrane – a thin, flexible barrier made of fat molecules that separates the inside of the cell from everything outside it. This membrane is selective by design. It keeps some things in and most things out.
But a cell that was truly sealed off from the world would die quickly. It needs water. It needs nutrients. It needs to expel waste. And in plants, cells also need to exchange gases – pulling in carbon dioxide for photosynthesis and releasing oxygen as a byproduct.
Getting all of this across a membrane that is, by nature, resistant to most molecules is the fundamental logistics challenge of all cellular life. Aquaporins are one of biology’s most elegant solutions to it.
An aquaporin is a protein that sits embedded in the cell membrane, forming a narrow, water-filled tunnel just wide enough for specific molecules to pass through in single file. These tunnels aren’t passive holes – they’re sophisticated gates with built-in molecular filters that determine, with extraordinary precision, exactly what is allowed through. Some aquaporins pass only water. Others allow carbon dioxide to diffuse through, speeding up the supply to photosynthetic machinery. Some are specialized for boron, a mineral critical to cell wall construction. Others can transport hydrogen peroxide – a molecule that, in the right concentrations, acts as a chemical messenger carrying signals about stress and damage throughout the plant.
A single plant cell may have thousands of aquaporins embedded in its membranes. Multiply that across billions of cells, add in the fact that the plant can dial the activity of each type up or down depending on its needs, and you start to understand why these proteins matter so much. They are not background infrastructure. They are active decision-makers in the plant’s moment-to-moment operation.
Forty-Eight Valves
The onion genome – one of the largest and most complex among cultivated plants, sprawling across roughly 16 gigabases of DNA – contains 48 distinct aquaporin genes. Each produces a slightly different protein with a slightly different job. Together they form the plant’s complete internal plumbing system.
They fall into four families, each stationed in a different part of the cell.
The PIPs (Plasma membrane Intrinsic Proteins) are the outer-wall specialists. Fifteen of them, embedded in the membrane that borders the cell’s exterior, handling the critical business of pulling water in from outside and letting carbon dioxide diffuse through to where photosynthesis happens.
The TIPs (Tonoplast Intrinsic Proteins) are the largest family at nineteen members, and they work the membrane surrounding the vacuole – a large fluid-filled compartment that takes up much of the interior of a mature plant cell. The vacuole is the cell’s reservoir and recycling center, and the TIPs manage the flow of water, nitrogen compounds, and other molecules in and out of it.
The NIPs (Nodulin 26-like Intrinsic Proteins) are a smaller, more specialized group of eight. Originally studied for their role in a completely different context – nitrogen-fixing partnerships between legumes and soil bacteria – the onion’s NIP proteins have been repurposed for a different set of tasks: moving minerals like boron and silicon across membranes, and shuttling hydrogen peroxide to where it needs to go.
The SIPs (Small Basic Intrinsic Proteins) are the quietest family, six members working inside the cell’s internal manufacturing network – the endoplasmic reticulum – on housekeeping tasks that maintain basic stability.
Each aquaporin’s specific job is determined by the precise arrangement of a handful of amino acids deep inside its tunnel. Tiny differences in these arrangements – sometimes just a single amino acid substitution – can mean the difference between a channel that only passes water and one that also passes carbon dioxide, or one that’s selective for boron over everything else. Researchers can now read these molecular signatures and predict, with reasonable confidence, what any given aquaporin transports without having to test it directly.
What’s particularly striking about the onion’s aquaporin family is that two subfamilies – the PIP1s and the TIP2s – appear to have been duplicated multiple times during the onion’s evolutionary history, generating expanded sets of closely related but subtly different proteins. This isn’t random. It’s functional flexibility. Having ten slightly different versions of a water channel allows the plant to deploy different variants in different tissues, at different times, under different conditions – turning what would be a blunt instrument into a precision tool.
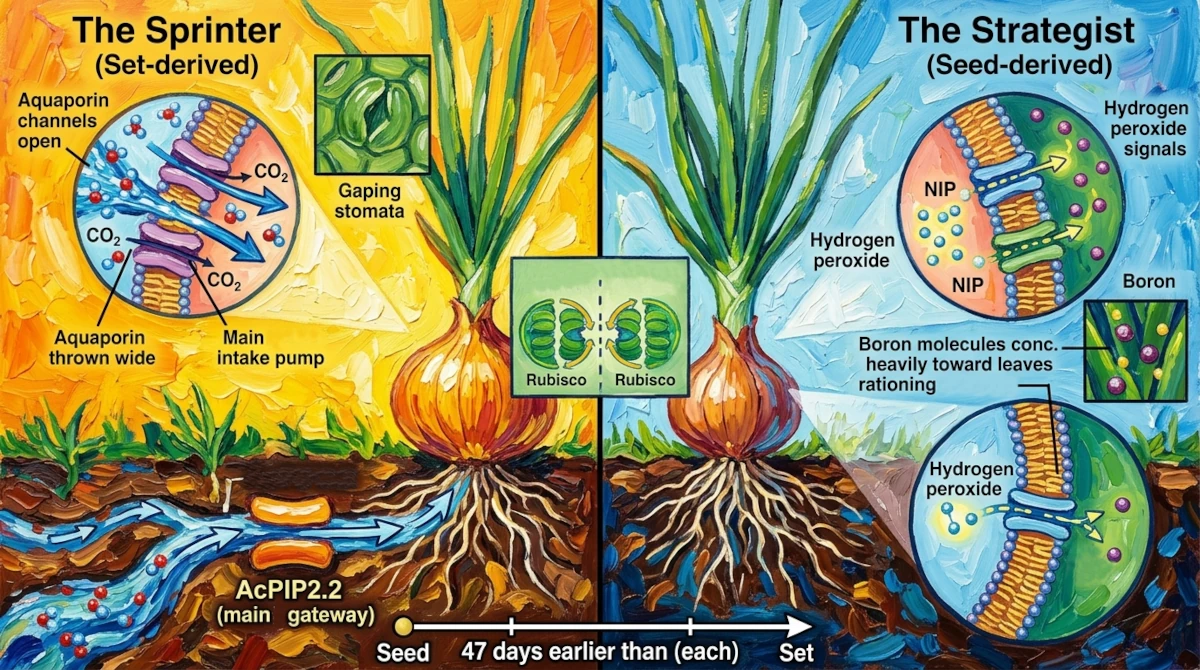
The Sprinter and the Strategist
Now we can return to those two plants growing side by side, and to the molecular story that separates them.
What the set-derived plant is doing
The onion set is a plant with a past. It was grown from seed, allowed to develop for part of a season, then harvested young, dried, and put into storage. What remains is a small, dormant bulb packed with stored energy – carbohydrates, nutrients, a pre-formed vascular architecture waiting to be reactivated.
When you plant a set and it breaks dormancy, the plant doesn’t ease into growth. It launches. The stored reserves provide immediate fuel, and the plant’s molecular machinery – as if responding to a starter’s gun – shifts into a configuration optimized for speed.
The effect on gas exchange is dramatic. Set-derived plants open their stomata (the tiny pores on their leaf surfaces through which gas exchange happens) wider and keep them open longer. They take in more carbon dioxide, release more water vapor, and fix more carbon through photosynthesis per unit of time. Their internal CO₂ levels actually run lower than those of seed-derived plants, not because they’re taking in less, but because they’re burning through it so fast that photosynthesis consumes it almost as quickly as it arrives.
To sustain this metabolic pace, the plant deploys a specific aquaporin configuration. In the roots, a protein called AcPIP2.2 becomes the dominant water-uptake channel – essentially the main intake pump for the whole hydraulic system. In the leaves, AcPIP1.1 and several TIP2 family members are elevated, maintaining the high-throughput plumbing that rapid photosynthesis and growth demand.
There’s also a clever molecular trick that set-derived plants use to fine-tune the balance between water and CO₂ transport. PIP1 and PIP2 proteins can physically associate with each other, forming joint structures. When they do, the PIP1 proteins – which would otherwise be stuck in internal cell compartments – get escorted to the outer membrane where they can assist with CO₂ diffusion. By adjusting the ratio of these joint structures, the plant can calibrate precisely how much of its membrane capacity is devoted to water versus gas transport, tuning its internal plumbing to the demands of the moment.
The result: a plant that grows fast, acquires carbon aggressively, and accumulates biomass at a rate that seed-derived plants simply cannot match.
What the seed-derived plant is doing
The seed has nothing to fall back on. A few days after germination, a seed-derived onion seedling is a tiny, fragile thing with shallow roots and barely any leaf area. Its world is full of threats: late frosts, dry spells, competition from weeds, soil crusts that block emergence. Growing fast in this situation is less important than surviving.
The molecular response to this situation is systematic caution.
Across all tissues – leaves, roots, and the developing bulb – seed-derived plants maintain elevated levels of hydrogen peroxide. This might seem alarming; hydrogen peroxide is, in concentrated form, a corrosive compound. But at the low, controlled levels found in plant tissues, it functions as a signaling molecule. It’s a chemical message that says: stay alert. Keep the defenses active. Don’t let your guard down.
The result is a plant in a state of what researchers call “physiological alertness” – its antioxidant systems are primed, its stress-response pathways are engaged, and its development proceeds more cautiously as a consequence. It grows more slowly, but it’s harder to kill.
The distribution of boron tells a similar story. Boron is essential for building and maintaining cell walls – the rigid structures that give plant cells their shape and strength. A plant running low on boron risks structural failure during rapid growth. Seed-derived plants handle this risk by concentrating boron heavily in their leaves (where the most vulnerable new growth is happening) while keeping levels relatively low in roots and bulbs. It’s deliberate rationing: protect the most critical tissues first, even if it means the rest of the plant operates on leaner supplies.
The aquaporin signature behind all of this is dominated by NIPs. Several NIP family members – AcNIP1.1, AcNIP3.1, AcNIP5.1, and AcNIP5.2 – are prominently active in the roots of seed-derived plants, handling the redistribution of boron and hydrogen peroxide in ways that serve the plant’s conservative resource management strategy. Two PIPs, AcPIP1.2 and AcPIP1.4, are elevated as well, associated with steady, stable water transport rather than high-throughput flux. Where the set-derived plant has a turbocharger, the seed-derived plant has a governor.
The Most Surprising Finding
Given all of the above, you might assume that set-derived plants simply have better photosynthetic machinery – more efficient enzymes, more productive carbon-fixing proteins. The sprinter, in this analogy, would have a faster engine.
But that’s not what the data shows. When researchers measured the efficiency of the core photosynthetic enzyme (Rubisco) and the ratio of carbon gained to water lost, they found no meaningful difference between the two types of plant. The biochemistry is identical. The engine is exactly the same.
The only difference is how much fuel – in the form of CO₂ – the aquaporin network delivers to that engine.
Set-derived plants don’t outperform seed-derived plants because their cellular machinery is superior. They outperform them because their plumbing removes a bottleneck. By upregulating aquaporins that improve CO₂ diffusion through leaf tissues, they ensure that the photosynthetic machinery is never waiting for carbon to arrive. The limiting factor in seed-derived plants isn’t biochemical – it’s hydraulic.
This is an important finding because it suggests that the onion’s growth potential isn’t constrained by its fundamental metabolism. The ceiling is higher than the seed-derived plant is reaching. And the reason it doesn’t reach that ceiling isn’t a flaw in its enzymes – it’s a choice embedded in its aquaporin network.
Forty-Seven Days
The most striking demonstration of the high-flux strategy in action is something researchers call the temporal advantage.
In experimental conditions, onion sets planted 47 days later than seeds still reached the same developmental milestone – the three-to-four true-leaf stage, an important marker of seedling establishment – at the same time. The sets, starting nearly seven weeks behind, sprinted hard enough to close the gap entirely.
Forty-seven days. That’s not a small difference. That’s almost a month and a half of calendar time erased by a difference in aquaporin gene expression.
Why This Matters Beyond Onions
For farmers and plant breeders, this research opens up a set of concrete possibilities.
The identification of specific aquaporins with specific roles gives breeders something precise to work with. AcPIP2.2, the dominant root water-uptake channel in set-derived plants, becomes a candidate target for selecting high-vigor cultivars – plants that inherit the hydraulic capacity of set-derived growth even when started from seed. AcNIP5.1, the key boron transporter in seed-derived plants, could be targeted to address the nutrient distribution problems that make seed-propagated crops vulnerable. AcPIP1.3 and AcPIP2.5, identified as CO₂-transport aquaporins, offer a route to improving photosynthetic efficiency by reducing the internal resistance that slows CO₂ diffusion in slower-growing plants.
The broader implication is that propagation mode is not just an agronomic detail – it’s a molecular dial. Turn it one way and you get a plant optimized for speed. Turn it the other way and you get one optimized for resilience. Understanding which aquaporins control which dial means, in principle, that future varieties could be designed to sit anywhere on that spectrum, tuned for the specific demands of a particular climate, season, or farming system.
Reading the Onion Differently
There’s something quietly wonderful about what this research reveals.
The onion is not a complicated plant in the way that a rainforest tree or a coral reef organism is complicated. It doesn’t have a brain, a nervous system, or anything we’d recognize as deliberate decision-making. And yet, embedded in the membranes of its cells, it carries a system of molecular gates so sophisticated that it can essentially read its own origin story – seed or set – and respond by reprogramming its entire transport network accordingly.
It doesn’t think about this. It doesn’t choose. The molecular response to being a set-derived plant or a seed-derived plant unfolds automatically, driven by gene expression patterns established in the earliest moments of growth. But the outcome is as if the plant had made a strategic decision: sprint, or conserve. Open the throttle, or stay alert.
Two onion plants, side by side in the same field, living out two completely different biological philosophies – both written in the language of forty-eight tiny proteins embedded in their cell membranes.
The humble onion, it turns out, contains multitudes.
Think you’ve mastered it? Test yourself with the 5-question quiz below – it only takes a minute.
Quiz: Can You Think Like an Onion?

Source
Study: Propagation mode shapes contrasting growth strategies through aquaporin networks in onion
Authors: Gloria Bárzana, Micaela Carvajal (2026)
Read the full paper: https://www.biorxiv.org/content/10.64898/2026.05.29.728701v1






Leave a Reply